
抗炎是抗衰老的一种策略!
快速发展的衰老研究或老年科学领域的科学家普遍认为,我们可以活得更久。少数化合物已被证明可以略微延长实验动物的寿命,但一些科学家更加雄心勃勃。英国伯明翰大学炎症与衰老研究所分子生物老年学教授João Pedro de Magalhães认为,人类可以活1000岁。他仔细研究了非常长寿的动物的基因组,如弓头鲸(200岁)和裸鼹鼠。他得出的令人惊讶的结论是:如果我们在细胞水平上消除衰老,人类可以活一千年,甚至可能长达2万年。这倒是符合中国人曾经的一种称呼,千岁和万岁。João Pedro de Magalhães长期从事衰老生物学研究,发表了系列研究论文,利用人工智能和现代生物医学研究手段,对抗衰老提出了独特的观点。这里我们介绍他10年前写的一篇论文,该文章对衰老提出了一种看法,认为衰老和炎症的关系密切。今天这一观点仍然具有一定地位,有一些研究也支持这一看法。初步研究发现,氢气对老年人各种衰老相关指标有改善作用,特别是炎症相关指标,显示出氢气对抗衰老的潜在价值。我们对比关于炎症在衰老中的地位,结合大量氢气抗炎症,尤其是慢性代谢炎症的作用证据。可以推测,氢气通过抗炎症实现抗衰老的价值非常高。又由于氢气对人的安全性高,适合长期使用,这非常符合抗衰老需要满功夫出细活的策略。Cell resilience in species life spans: a link to inflammation? - Finch - 2010 - Aging Cell - Wiley Online Library物种寿命的差异归因于各种应激源期间的细胞存活,这里称为“细胞恢复力”。在原代成纤维细胞培养物中,暴露于自由基、低血糖、体温过高和各种毒素期间的细胞恢复力与鸟类和哺乳动物的物种特征寿命显示出普遍一致的相关性。然而,成纤维细胞培养物中细胞恢复力与不同物种寿命的机制联系知之甚少。我们提出,某些实验应激源与先天免疫炎症反应期间体内体细胞损伤有关,特别是对ROS,低葡萄糖和高热的抵抗力。根据这一假设,体细胞恢复力决定了物种在自然环境中反复感染和创伤期间的寿命差异。感染和损伤使局部成纤维细胞和其他细胞暴露于巨噬细胞产生的ROS和局部温度升高。全身性,急性期免疫反应引起低血糖和高热。我们认为,细胞对炎症中发生的体细胞应激源的恢复力在长寿的进化中很重要,并且长寿物种对免疫相关应激源的抵抗力特别强。这一假设进一步阐明了柯克伍德的一次性体细胞理论。我们建议将用于比较研究的压力源和标志物的电池扩展到与宿主防御及其生态特异性相关的其他细胞类型和附加参数。介绍我们提出了一个统一的假设,即成纤维细胞对应激源的恢复力的物种差异与宿主防御中的先天免疫机制的物种寿命有关。原代成纤维细胞培养物广泛用于评估细胞对各种应激源(包括活性氧 (ROS)、高热和低血糖)的恢复力与物种寿命的相关性,总结如下:表1.这些研究通常显示物种寿命与短期体外成纤维细胞存活和修复能力呈正相关。然而,目前尚不清楚细胞对各种压力源的恢复力如何与衰老和成人死亡的原因有关。
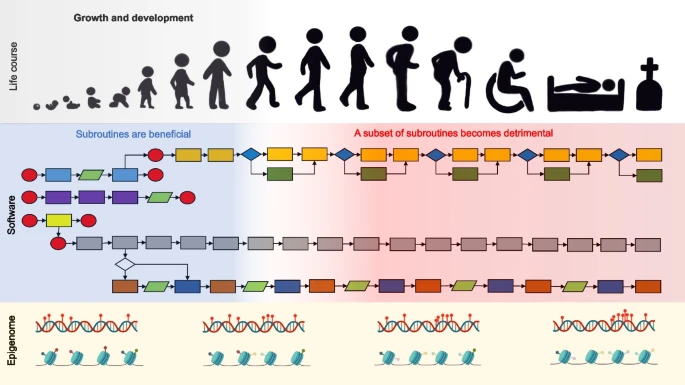
历史上以哈曼的自由基衰老理论为指导,即ROS损伤和氧化应激是与年龄相关的功能障碍和疾病的主要原因。显然,在受保护的人类和家畜种群中,晚期衰老期间死亡的主要原因涉及氧化损伤,例如动脉粥样硬化,肿瘤形成,糖尿病,肥胖和神经变性。然而,在自然种群中,捕食、感染和创伤是所有年龄段死亡的主要原因。例如,在野生黑猩猩中,感染导致大多数(67%)成年死亡(Williams等人,2008年;古道尔,1986,芬奇2010)。直到最近,在历史上的欧洲人口中,感染也导致了所有年龄段的大多数死亡(Preston,1976年),以及获得有效药物的机会有限的狩猎采集者(Hawkes等人,2009年;芬奇,2010)。尽管在寿命较短的物种的自然种群中几乎没有关于老年特异性病理学的记录,但大多数成年人似乎不太可能存活足够长的时间,以致于引起与炎症和氧化损伤相关的人类衰老的慢性动脉和脑部疾病。之前关于比较抗逆性的工作主要集中在对长寿动物的需求上,以限制氧化和其他损伤。这种关注忽略了很大一部分损伤是由感染和感染相关的组织反应引起的的可能性。我们认为细胞对ROS,高热和低血糖(表1)与急性感染和创伤性组织损伤以及慢性局部炎症期间的宿主体细胞损伤高度相关。具体来说,细胞对ROS,热应激和低葡萄糖的恢复力是先天免疫的急性期炎症反应的基础,其涉及巨噬细胞产生ROS,体温过高(发烧)和低血糖。因此,组织成纤维细胞和其他细胞作为旁观者暴露于类似的应激源,导致氧化损伤,用于比较实验老年学。炎症期间的氧化损伤,作为衰老的普遍机制,也在Kirkwood的一次性体细胞衰老理论中得到认可,该理论认为在生殖能量分配和各种分子损伤的体细胞修复之间进行权衡的重要性(Kirkwood,2005;德雷诺斯和柯克伍德,2005 年)。最后,我们建议额外的压力源和细胞弹性的测量以及与物种比较的生态相关性标准。先天免疫的急性期反应先天免疫的急性期反应使宿主能够对可传播的病原体和伤口进行短期防御。B细胞和T细胞对入侵病原体抗原的适应性免疫反应增强了随后的存活率。感染和伤口产生局部和全身反应,从而有效地修复组织并防止化脓性感染。复杂的炎症过程仍然恰如其分地体现在塞尔苏斯的经典格言中:rubor et tumor cum calore et dolore(发红和肿胀伴热和疼痛)。许多急性期反应也持续存在于与人类衰老疾病相关的慢性炎症中。炎症导致氧化应激和DNA损伤哈曼理论中描述的自由基也是宿主防御的基础。微生物病原体可激活血巨噬细胞和中性粒细胞,通过NADPH氧化酶产生呼吸爆发,产生超氧化物(O2−)并增强吞噬作用。在中性粒细胞中,细菌杀伤发生在吞噬细胞液泡中,通过NADPH氧化酶驱动的pH改变,而不是由ROS(Segal 2005)。源自超氧化物的其他ROS包括由H202和过氧亚硝酸盐(ONOO−) 和 O2−.组织损伤迅速诱导来自常驻局部细胞和招募白细胞的ROS(Sen和Roy2008)。在斑马鱼幼虫无菌损伤模型中,H2O2从上皮细胞募集白细胞到伤口边缘(Niethammer等人2009)。宿主防御过程中产生的ROS会对旁观细胞和细胞外分子造成氧化损伤。在肿瘤发生的两阶段模型中,慢性炎症被认为是肿瘤启动子(Coussens和Werb 2002)。炎症引起的ROS损伤的一个主要例子与胃癌有关,胃癌是全球恶性肿瘤中排名第二的死因(Herrera等人,2005年;世卫组织,2009年)。胃癌的一个主要原因是幽门螺杆菌的慢性感染:大约15%的携带者发展为消化性溃疡,1%进展为癌症(幽门螺杆菌和癌症合作小组,2001)。一个主要的致癌途径涉及局部炎症细胞对细胞外幽门螺杆菌的反应。浸润巨噬细胞和中性粒细胞产生ROS,破坏相邻上皮细胞(8-OHdG)中的DNA(Farinati等人,2003)。同时,端粒DNA的缩短与粘膜细胞增殖有关(Kuniyasu等人,2003)。抑制COX-2的抗炎非甾体抗炎药大大减少了人群中的胃癌(Wang等人,2003)。克罗恩病和其他独立于幽门螺杆菌的特发性肠道炎症性疾病也会增加患胃癌的风险(Itzkowitz和Yio,2004)。引起氧化应激和DNA损伤的炎症的旁观者效应延伸到许多其他炎症状况(Meira 等人,2008 年;川西等人,2006;深田和阿布雷乌,2008年)。因此,成纤维细胞恢复力(表1)可被视为炎症期间ROS旁观者损伤的模型。选择抗菌热疗可产生耐热成纤维细胞发烧是宿主防御的关键部分,进化为抑制微生物病原体在其最佳温度以上的生长;甚至荸荠也会出现发烧(Hasday等人,2000年;内斯和威廉姆斯1996)。然而,热疗可以通过半胱天冬酶-2008和-3诱导细胞死亡(例如Bettaieb和Averill-Bates,9)(Nagarsekar等人,2008)。热热诱导的细胞凋亡与焦亡重叠,焦亡是一种炎症期间的新型细胞死亡,是半胱天冬酶-1依赖性的(Bergsbaken等人,2009)。除了全身性高热之外,有据可查但鲜为人知的是,局部炎症也会增加局部温度,正如塞尔苏斯的格言一样。最容易理解的例子是晚期动脉粥样硬化斑块中的热点,在相邻动脉表面上方高达3°C,并且与局部巨噬细胞密度相关(Madjid等人,2002年;谭和唇 2008;图图扎斯等人,2007年)。局部热疗是通过诱导UCP2产生的,UCP2008将线粒体ATP的产生与呼吸分离,从而产生辐射能(Van De Parre等人,2009)。我们认为局部热疗可能通过抑制与受损血管组织相关的局部细菌来适应。肺炎衣原体和其他常见细菌在动脉粥样硬化斑块中检测到,并被争论为血管事件的危险因素指标(Di Pietro等人,2008 年;斯塔森等人,2000年;卡拉约古等人,2004年)。这些机制可能延伸到手足骨折炎症期间观察到的局部皮肤热疗(Huygen等人,2)。由于成纤维细胞表达UCP2008(Mori等人,2),我们建议将UCP1998诱导和产热包括在物种比较中。由热疗诱导的其他标志物可能包括细胞ROS的水平(Flanagan等人,1)以及与细胞耐热性相关的SOD2和SOD1985活性(Loven等人,2002;星田等 2000;山下等人,<>年)。低血糖和细胞抵抗先天免疫急性期的低血糖,例如小鼠LPS注射后3-6小时(Oguri等人,2002;阿里等人,2008年;Cryer et al 2009)被理解为一种抑菌机制。在比较研究中(表1),来自长寿侏儒小鼠的成纤维细胞显示出对葡萄糖剥夺诱导的代谢标志物变化(WST-1减少)的抵抗力增强,这与对细胞毒性物质(如过氧化氢)的抵抗力呈正相关(Leiser等人,2006)。这些研究中的葡萄糖饥饿水平(0.1-0.4毫克/毫升)处于扰乱人类认知和行为的短暂性低血糖的低临床范围(<0.50毫克/毫升,或<50毫克/分升,Cryer 2007;Cryer et al 2009)。成纤维细胞的急性期恢复假说和相关性我们认为,细胞对免疫相关应激源氧化损伤的恢复力是长寿系统的关键组成部分。感染由摄入和吸入可传播的病毒和细菌病原体以及体外寄生虫引起。感染性炎症负荷对寿命的重要性体现在畜牧业改进导致实验室啮齿动物寿命延长,最大限度地减少了慢性肺部疾病、体外寄生虫和其他地方性感染:对于杰克逊实验室的 C57BL/6J 小鼠,寿命从 50 年(1957 个月)到 17 年(1970 个月)增加了 >28%(Finch 2007,第 137 页)。成纤维细胞,作为物种比较中的主要测试细胞(表1),与皮肤在达尔文世界中的重要性有关。皮肤成纤维细胞普遍受到体外寄生虫和微生物动物群的慢性炎症,以及战斗期间受伤或侵略性社会梳理引起的慢性炎症(Takahashi和ori,1982;利特文等人,2007年)。寿命很长的裸鼹鼠在殖民地内攻击期间会受到深深的咬伤,刺穿内脏,偶尔会致命(Lacey等人,1991年;克拉克斯和福克斯,2001年;克里斯·克拉克)。与咬伤相关的脓肿含有巴斯德菌属和嗜血杆菌属(Artwohl等人,2002 年)。长寿裸鼹鼠对反复伤害的恢复力支持一次性体细胞假说(Kirkwood 2005),即选择体细胞恢复力可以延缓衰老。皮肤损伤刺激反应可去除坏死碎片并防止使用许多先天免疫系统基因的感染;成纤维细胞可能被刺激增殖并经历细胞凋亡(Fujiwara等人,2007;圣地亚哥等人,2004年)。损伤诱发 H2O2如上所述,上皮细胞的产生似乎引导白细胞的流入(Niethammer等人,2009年)。SOD-1和-2以及过氧化氢酶的诱导(Steiling等人,1999;舍费尔和维尔纳2008)与 H 的清除有关2O2和其他 ROS,这会损害伤口修复(Wasserbauer 等人,2008 年;威尔格斯等人,2005年)。虽然成纤维细胞单层可以用作损伤模型,但我们不知道与物种寿命相关的比较研究。除成纤维细胞外,其他细胞显示出寿命损伤的反比关系。暴露于氧化应激(高血糖症)的主动脉内皮细胞在实验室小鼠(C57BL / 6)中比长寿鹿小鼠(Peromyscus leucopus)显示出更多的炎症基因诱导(Labinskyy等人,2009)。同样,白细胞主动脉细胞产生的线粒体ROS少于实验室小鼠,与其他比较一致(表1).这些物种寿命与线粒体ROS和动脉组织中细胞对ROS的脆弱性呈负关系的发现与成纤维细胞比较一致,并为在比较研究中扩大体细胞类型的范围提供了基础。粘膜上皮细胞在未来的物种比较中值得考虑,因为它们在肠道和口腔防御中起着关键作用。免疫和细胞死亡基因序列的基因组比较在理解成纤维细胞表型的物种差异如何与DNA序列和基因表达方面的基因组差异相关方面存在重大差距。在编码序列水平上,免疫反应基因显示出许多相关的物种差异;有迹象表明,哺乳动物的谱系特异性创新会减弱长寿物种的先天免疫反应(Wang等人,2006年)。来自同义突变与非同义突变的密码子比率统计的免疫反应基因的正选择已被充分记录(Nielsen 等人,2005 年)。自人类与黑猩猩分道扬镳以来,与细胞凋亡相关的基因的物种差异占正选择基因的相当一部分(Nielsen et al 2005)。除了密码子的变化外,基因缺失和表达谱的变化也存在许多物种差异。人类和黑猩猩基因组显示出广泛的基因缺失(Olson 1999;Wang等人,2006),其中许多与炎症反应有关,由与免疫功能相关的示例说明。黑猩猩有ICEBERG,IL1F7和IL1F8的缺失,它们处于抑制半胱天冬酶-1表达的途径上(Osborn等人,2005);这些缺失可能会增加对LPS诱导的IL-1的反应(Humke等人,2000)。人类与类人猿的不同之处在于缺乏N-乙醇酰神经氨酸(Neu5Gc)(Varki 2009),这是由于CMAH失活,CMAH酶促地将Neu5Ac转化为Neu5Gc。CMAH在0万年前的失活对靶向Neu5Ac的人类特异性病原体有影响:黑猩猩疟疾寄生虫在红细胞入侵期间结合Neu5Gc,而人类恶性疟原虫结合Neu5Ac。对人类和黑猩猩的比较表明,与炎症反应和细胞增殖相关的基因更有可能显示基因丢失或获得,例如APOL5仅存在于人类中并介导锥虫抗性(Perry等人,1)。来自不同物种的成纤维细胞的表达谱相对不发达。我们建议,具有不同寿命的物种之间的基因表达比较可能会揭示与衰老相关表型有关的基因。特别是物种特异性细胞对压力和炎症的反应可能有助于长寿的进化。应使用下一代测序技术,该技术允许深度RNA测序,其表达谱分析比传统微阵列更敏感(de Magalhaes等人,2010)。这些细胞反应在啮齿动物中的基因表达谱可以识别在长寿啮齿动物中进化出独特作用的基因和途径,并可能有助于它们的寿命。例如,啮齿动物与人类的比较显示,成纤维细胞模型中与ROS相关的DNA修复基因存在差异:紫外线损伤DNA结合蛋白(UV-DDB)。它在小鼠和仓鼠表皮细胞中的表达量要低得多,这与相对于人类的全球基因组修复缺陷一致。DDB2的异位表达增加了小鼠成纤维细胞的DNA修复,并减少了体内紫外线诱导的皮肤癌(Alekseev等人,2005)。我们预计未来的研究将使用新的转录分析技术揭示与长寿相关的基因表达模式的其他物种特异性差异。对于人类与灵长类动物的比较,最详细的研究可能是Affymetrix GENECHIP 5.0对成年人类,倭黑猩猩,黑猩猩和大猩猩的成纤维细胞的比较,并通过北方印迹证实了选择的变化(Karaman等人,2003):在区分的10,000个基因中,约1%显示出≥2倍的物种差异,包括与宿主防御的各种关联:人类HLA-E的表达更高, MICA,SDF1和TGFβ1,但glypican 3(一种肝素硫酸蛋白多糖)的表达要低得多。另一个与免疫功能直接相关的基因是糖蛋白siglec-5在人类T细胞中的表达非常低,这是人类与黑猩猩CD4 T细胞免疫反应性更大的一个因素,如表达操作所示(Nguyen等人2006)。与癌症的进化有关,erbB原癌基因在人类和两只类人猿中与几种寿命较短的灵长类动物相比具有更高的表达,而myc,ras-K和src没有差异(Nakamura和Hart,1987)。在物种比较中,这些基因都没有在成纤维细胞恢复力的背景下被考虑过。这些差异表明免疫反应发生了重大的进化变化,这是人类长寿生物学的一部分。很明显,无论免疫反应的微观进化如何,都没有单一的测定可以解决物种差异的多个方面。炎症引起的体细胞损伤与一次性体细胞衰老理论有关,根据该理论,寿命由积累体细胞损伤的速率决定,该速率被修复和再生的能量分配水平所抵消(Kirkwood 2005)。因此,与单胎出生5年或更长时间的长寿黑猩猩相比,每次交配有多个后代的短命小鼠在修复氧化损伤方面的投入更少。这可以很容易地在单细胞生物(如细菌或酵母)中观察到,它们在进入非生殖阶段(后双相和固定阶段)后对多种胁迫的抵抗力要强得多。这些物种差异的基因组基础可以通过RNA分析和DNA序列比较来解决。我们预计,防止炎症损伤和介导修复的基因的物种差异将为长寿物种的修复和再生机制提供线索,这可能解释长寿的进化。系统发育和生态问题Kapahi等人(1999年)的研究是一个基准,因为它包括了包括人类在内的四个分类学目中的哺乳动物物种。对8种啮齿动物的研究通常证实了细胞恢复力 - 寿命关系(Harper等人,2007)。最近对物种进行体外比较的研究扩大了分类单元中代表的寿命范围。将超长寿裸鼹鼠(Heterocephalus glaber,寿命>28 年)包括在内,相对于实验室啮齿动物,现在的寿命范围是 7 倍(Gorbunova 等人,2008 年;布芬斯坦2008)。除Heterocephalus glaber外,本研究的其他物种都是陆生的。鼹鼠的加入引发了关于实验压力源的生态相关性的新问题:紫外线照射似乎与数百万年来一直避开阳光的鼹鼠无关。其他生态生理适应可能对实验设计很重要。H. glaber完全生活在长长的地下洞穴中,这个生态位与其他深海动物一起占据了至少20万年(早期中新世)(Nevo等人,1999年)。H. glaber的核心温度较低<34°C,被认为接近极热(Buffenstein和Yahav,1991)。尽管如此,在先天免疫激活期间可能会出现核心温度升高的发热(R. Buffenstein, pers. comm.)。低核心温度在培养条件下引入了物种特异性因素。与陆生啮齿动物的这些主要生态差异可能是几种异常的一个因素。光头嗜血杆菌成纤维细胞对 过氧化氢 ,UV,鱼藤酮和低葡萄糖更敏感,而小鼠在心脏线粒体产生的ROS方面与两只鼹鼠没有差异(Lambert等人,2007)或肝脏抗氧化剂SOD,过氧化氢酶和谷胱甘肽的水平(Andziak和Buffenstein 2006)。此外,在体内,H. glaber具有比小鼠更高的氧化脂质和蛋白质负荷(Andziak等人,2006)。异脑脂质对氧化损伤过氧化氢的敏感性更高被认为是“...挑衅,因为它不支持大多数氧化应激和长寿模型“(Salmon等人,2008)。然而,H glaber的栖息地与其他啮齿动物非常不同:其狭长隧道中的气体成分具有非常低的pO2(6%,缺氧)和高 pCO2(10%,高碳酸血症),这将杀死大多数啮齿动物(van Aardt et al 2007)。相应的适应包括具有更高氧亲和力的血红蛋白和改变的酸碱调节(更大的玻尔效应)(Johansen等人,1976;van Aardt et al 2007),我们建议这可以扩展到介导应激反应的其他生化系统的氧化还原敏感性。其他寿命为 20 年的啮齿动物包括海狸、豪猪和松鼠(Gorbunova 等人,2008 年;奥斯塔德 2009);其他鼹鼠的寿命可能较短,但观察到的少量排除了确切的结论(http://genomics.senescence.info/species/)。对未来研究的建议由于急性期反应使细胞同时暴露于高热和低血糖,因此未来的比较可以考虑热休克反应(例如HSP70-1,2),产生局部热疗的线粒体UCP2以及葡萄糖调节和转运蛋白(例如Grp58,GLUT-4)。热和低血糖的组合应通过深度RNA分析进行检查。我们假设寿命将随着体细胞恢复力而扩展,以适应宿主防御期间经历的压力源组合。细菌内毒素也可用作探针。常见革兰氏阴性菌感染的LPS内毒素诱导成纤维细胞中的炎症基因诱导(Perfetto等人,2003;华纳等人,2004)。SOD-2 mRNA的LPS诱导(Visner等人,1990)增强了对H的抵抗力2O2 (罗尔等人,2008年)。预计基底细胞和应激细胞中的深层RNA分析将进一步获得见解。在正在进行的啮齿动物研究的同时,我们鼓励进一步比较人类细胞与类人猿和其他表达数据非常有限的类人猿。另一个考虑因素是用于比较的人类细胞中的等位基因变异,据我们所知,在物种比较中尚未考虑这一点。例如,半胱天冬酶12在人群之间假基因水平上有所不同,这增加了对败血症的抵抗力(Wang等人,2006);活性等位基因(Casp12-L)虽然在大多数亚洲和欧洲人群中很少见(<1%),但在撒哈拉以南非洲很常见,高达60%(Kachapati等人,2006年);黑猩猩和其他哺乳动物有活跃的Casp12。除了与感染有关外,半胱天冬酶12还与淀粉样蛋白诱导的神经元凋亡有关,这可能与黑猩猩和其他类人猿明显不存在阿尔茨海默病有关(Finch 2010;黑猩猩测序和分析联盟,2005年)。另一个相关的等位基因系统是apoE,它影响细胞因子分泌(Vitek等人,2009),以及寿命,阿尔茨海默氏症和心脏病。主要的组织相容性基因复合物(Mhc)也具有与体细胞恢复力相关的等位基因变异。结束语体外细胞恢复力与物种寿命之间的联系是衰老生物学中一个迷人而重要的范式。我们认为,对多种类型炎症事件具有抵抗力的进化选择支持了一般观察,即长寿物种培养的细胞具有抗应激性。该假设进一步说明了Kirkwood的一次性体细胞理论与炎症(2005)相关的方面。我们预测,长寿物种对特定的免疫相关压力源具有更强的抵抗力,并基于这一假设提出了未来的研究。

(汇进升)采用物理方法生成活性氢原子活水,其特点是高浓度、弱碱性、小分子团和蕴含微量能量于一体的健康功能活水,能有效清除自由基,促进身体健康。
普通的氢气水产品中氢气浓度从01ppb到8ppb之间,氢化镁、镁、氢化钙和硅都是理论上可行的产氢气食品补充剂的活性成分。
汇进升活性氢原子水的氢气浓度达到了惊人的6000PPb,获得了国家专利,给人民大众带来了福音。

健康人群丨饮用建议
●将氢水当作日常饮用水;
●早晚各饮用400ml;
●日间饮用800-1200ml;
●40°的温水,小口多喝是最好的饮用方式。
选它☞:汇进升活性氢原子山泉水(氢浓度≥6000ppb)


【汇进升活性氢原子麦饭石山泉水】本着诚信、协同、创新的精神,积极寻找良好水源,经过对沂蒙山区多地实地勘察,终于在沂蒙山脉的食用水源保护地麦饭石矿岩区内的五彩山中发现了自涌山泉,该泉从沂蒙山脉岩层深处喷涌而出,富含硒、钙、锶、钾、钠、镁、偏硅酸等对人体有益的多种矿物质,经过高新技术制氢设备的激发,氢原子浓度稳定达到6000PPM以上,从而提高人体代谢能力,净化血液并修复细胞,提高人体自觉与自愈能力,是不可多得的健康饮用水。
常喝汇进升活性氢原子麦饭石山泉水,氢会进入肝、肾、胰岛腺、胃、小肠等部位,能调理身体机能,提高身体免疫力,对治病起很好的辅助作用和美容效果。【点击此链接后查看专家视频讲座】
- 上一篇:NIH长新冠临床试验公告 2023/8/22
- 下一篇:氢化珊瑚钙治疗代谢综合征患者临床研究 2023/8/22
